
Introduction
The British steel industry represents 26% of total UK greenhouse gas emissions, making it the 3rd largest industrial emitter of CO2 due to high energy consumption (Ambrosio-Albalá et al., 2023). Reducing energy consumption and CO2 emitted in the steel industry must be at the forefront if our nation is to become carbon neutral by 2050 (Knight and Walker, 2022).
My proposal creates an adapted biochemical cell, sharing similarities to a hydrogen fuel cell, for the reduction of iron oxides instead of the more commonly seen electricity production. Within its framework, the novel cell contains a membrane bound hydrogenases for the splitting of dihydrogen into its consecutive protons and electrons. Once split, the electrons are free to cross a synthetic membrane, facilitated by membrane bound MtrCAB proteins promoting the reduction of iron oxides.
The Hydrogenase and MtrCAB proteins in the biochemical cell will be examined thoroughly throughout this essay, giving insights into their structure and function. Equally important will be the reasonings and applications of the proteins’ integration into a practical biochemical cell (hereafter the ‘Sheffield Cell’).
Consequently, this biotechnology will result in pure iron as a product for industrial purposes, and water as a waste byproduct for further recycling as a coolant during iron manufacturing. The aim is to create an innovative product which reduces CO2 emissions and creates a circular economy.
Aim of the proposal
Releasing Fe3+ without producing CO2 emissions, and lowering the energy consumption needed, in the form of extreme heat, due to fossil fuel combustion.
6H2 → 12e− + 12H+
2Fe2O3 → 4Fe + 3O2 + 12e−
3O2+ 12H+ + 12e−→ 6H2O
Figure 1: Equations related to the Sheffield Cell
The Structure and Function of the Sheffield Cell
Hydrogenases are metalloenzymes catalysing the reaction of dihydrogen into 2 protons and 2 electrons (Lubitz et al., 2014). They have a diverse set of active sites, containing either [Fe] commonly observed within archaea, [FeFe] the only active site seen in eukaryotic hydrogenases, or [NiFe] found in archaea and prokaryotes such as cyanobacteria (Pathy et al., 2022). The [NiFe] hydrogenases in particular feature oxygen resilience characteristics within aerobic environments, making them a prime candidate for the Sheffield Cell in standard conditions (Shomura et al., 2011). The structure of [NiFe] hydrogenase constitutes a dimer protein, consisting of a large HoxG and smaller HoxK subunit (Lubitz et al., 2014). What is more, a cytochrome b is affiliated with the [NiFe] hydrogenase, aiding in the movement of electrons into the electron transport chain of the host bacteria – this model is represented by the MtrCAB complex for the electrons transport mechanism (Lubitz et al., 2014).
HoxG is the larger of the subunits responsible for the catalytic splitting of dihydrogen, inside the dinuclear [Ni-Fe] active site (Lubitz et al., 2014). Coordinately bound to its active site are CO and two CN- ligands, leaving a vacant catalytic site for H2 incorporation between the Ni and Fe (Lubitz et al., 2014). Further stabilisation is initiated by cysteines 80,84,546 & 549 of the Ni, and cysteines 84 & 549 coordinately bound to the Fe (Higuchi, et al., 1997), whilst additional proximal amino acids are nestled in the protein structure as follows: arginine, serine, glutamate, and a distal amino acid histidine (Fontecilla-Camps et al., 2007).
There are two contributing factors to the catalytic potential of the active site of [NiFe] hydrogenase. First, the central conformation of the [Ni-Fe] complex bound to dihydrogen forms a distorted unfavourable square planer conformation, due to the near 90-degree bond angle (Fritsch et al., 2011). Second, instrumental to the catalytic potential, the electron disparity experienced by the dihydrogen and active site complex lowers the pH of the active site below physiological conditions. The splitting of dihydrogen is potentially aided by like charge encounters of the protons from the dihydrogen and arginine (pKa 10, and therefore protonated), and unfavourable conformational environment of the active site/substrate complex. This results in steric hindrance and decreased molecular stability (Fontecilla-Camps et al., 2007). Once the splitting of dihydrogen has occurred and expulsion of protons from HoxG via the Mg2+ pathway has commenced, the electrons are free to pass further into the small subunit HoxK (Higuchi, et al., 1997). HoxK is the site of electron transport via the three [Fe-S] clusters (Fritsch et al., 2011), presented in the following order:
Proximal [4Fe-3S] coordinated by 6 cysteine residues, Cys 149, Cys 120, Cys 115, Cys 17, Cys 19, Cys 20
Medial [3Fe-4S] coordinated by 3 cysteine residues, Cys 230, Cys 249, Cys 252
Distal [4Fe-4S] coordinated by 3 cysteine residues and a histidine residue, Cys 215, Cys 221, Cys 190, His 187 (Fritsch et al., 2011)
Lastly, an important aspect of the HoxK is the carboxy terminal alpha helical secondary structure, acting as the embedment site to the host cell membrane, as well as the connection site for the cytochrome b electron carrier (Fontecilla-Camps et al., 2007). When deciphering between hydrogenases, the oxygen tolerance of [Ni-Fe], plus its natural position within the periplasmic membrane, offers promising applications during artificial membrane insertion. The oxygen tolerance is proposed, although not yet fully understood, to reflect the composition of the [4Fe-3S] structures within the small subunit of the enzyme (Shomura et al., 2011). The clusters differ from other standard hydrogenases containing a [4Fe-4S] in that they facilitate four coordinated cysteine amino acids, opposed to the Proximal [4Fe-3S] [NiFe] hydrogenase bound by six cysteines. The six cysteines further fabricate the distorted conformation of the Proximal [4Fe-3S], manifested by the additional Cys 120 & 19 (Fritsch et al., 2011). Furthermore, it has been suggested that the additional cysteines may aid in the oxygen tolerance within the membrane bound [NiFe] hydrogenase (MBH) (Fritsch et al., 2011).
MtrCAB is a Shewanella oneidensis membrane bound electron transport protein consisting of the subunits MtrC, MtrA and MtrB (Edwards et al., 2020). The MtrA unit is situated within the periplasmic region harbouring the decaheme c-type cytochromes matrix – accommodated by the beta barrel conformation of MtrB, further encased in the outer membrane (Edwards et al., 2020). The final unit MtrC protrudes into the outer membrane, hosting the terminal decaheme c-type cytochromes complex (Bretschger et al., 2007; Edwards et al., 2020). It has been suggested that either the heme 10 or 5 are responsible for the electron transfer to the extracellular substrate (Edwards et al., 2018), although the exact mechanism has not yet been finalised, the protein’s electron transport actions are known.
The Sheffield Cell consists of multiple membranes throughout the system, two of which will have proteins encompassed into the framework. Techniques such as electrical current destabilisation of the membrane, or the addition of a detergent for slow removal of biobeads are both favoured for protein insertion (Bruns and Kilbinger, 2017). Additionally, the construction of a porous membrane using a solid frame support for the insertion of MtrCAB may provide greater applicational success (Bruns and Kilbinger, 2017). Furthermore, the Langmuir-Blodgett & Langmuir-Schaefer (LB/LS) methods linking the ‘grafting to’ principle of polymerisation, involving a step-by-step process of chain precipitation, coinciding with the inclosing of MBH might be better suited (Bruns and Kilbinger, 2017). Regardless, each protein membrane insertion mechanism would need efficient experimentation to decipher the most desirable insertion technique.
For successful ion transfer and controlled reactions throughout the Sheffield cell, a proton-exchange electrolyte membrane and an oxygen permeable membrane preventing Fe3+ ion leakage would be required. Although still within its infancy, a possible proton-exchange membrane (PEM) could be synthesised using cellulose (Selyanchyn et al., 2020). Modifications would be necessary however, with additional functional groups such as sulfonation for an enhanced proton connectivity as currently cellulose is inefficient as a standalone material (Selyanchyn et al., 2020). Since cellulose is a renewable inexpensive polymer, replacements of current PEM’s using this renewable biomaterial could offer the optimum alternative for future applications. Oxygen permeable Perovskite oxides are an example of mixed ionic-electronic conducting membranes, which has attracted interest due to their competence in isolating oxygen (Zhu and Yang, 2019). Perovskite oxides have the formula ABO3 forging their lattice structure – A, accounting for alkaline earth/lanthanide metals, and B associated to the transition metals (Zhu and Yang, 2019). One limitation potentially hindering the system when utilising Perovskite oxides is its permeability to H2, and it would therefore require experimental simulations to prevent cross contamination (Zhu and Yang, 2019). Moreover, perovskite oxides operate at temperatures exceeding 700 ∘ C (Sunarso et al., 2017), which would ultimately disrupt additional components of the Sheffield Cell. To overcome these extreme operational temperatures, a recent study has investigated the use of dielectric barrier discharge plasma (DBD plasma), coupled to an oxygen permeable membrane (Zheng et al., 2022). The study applied an air plasma power of 15 W, allowing an activation energy reduction of the permeation process, leading to an increase in the oxygen flux by a factor approaching 30 and a temperature depletion of 600 ∘ C (Zheng et al., 2022). Despite this decrease in temperature achieved by maximising the DBD plasma, the use of oxygen permeable membranes still requires further innovation for uses at room temperatures.
Sheffield Cell Composition
The Sheffield Cell will be compartmentalised - dihydrogen upon entering the cell will interact with the larger subunit of the MBH, nestled on the outer side of its synthetic membrane. Once the MBH has split H2 into its successive protons and electrons, the protons due to repulsion are expelled back into the hydrogen containing compartment of the Sheffield Cell.
Further to this, the electrons move through the [4Fe3S] complexes within the small subunit of MBH. Drawing close, the electrons enter the MBH tail infixed within the membrane before joining the electron carrier cytochrome b. Here they are transported through MtrCAB by the heme network, bound within the adjacent synthetic membrane. The electrons can traverse the MtrCAB due to the lowering of the electrons’ energy in crossing the heme network, before passing into the site of iron oxide. The electrons now readily participate in the reduction of iron oxides as seen in Figure 1. Once reduction of iron oxide has occurred, both Fe3+ and oxygen are available to be extracted.
Reversing back, the positively charged H+ protons can freely cross the electrolyte membrane, readily reacting with O2 once released from the iron oxide. Oxygen diffuses into the H+ space via the oxygen permeable membrane producing H2O after reacting with 2H+, abandoning Fe3+in the system. As Fe3+ is a positively charged ion there are many options for its removal, in particular the addition of a chelating membrane or magnetism (Siekierka, Nowicka, and Ostrowska, 2023). Ultimately, the cell would feature two subunits fused during operations for the insertion of iron oxide and the removal of pure iron. The newly attained pure iron can then continue in the standard iron production line.
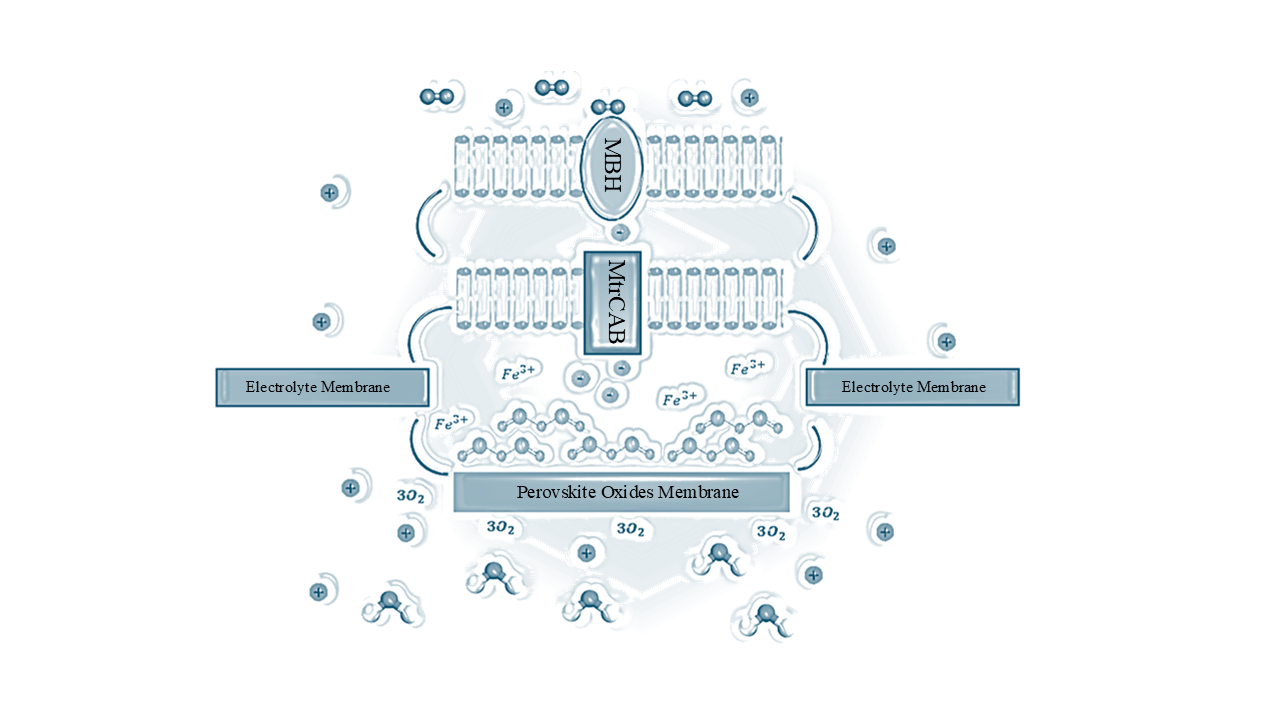 |
|---|
Figure 2. A representation of the Sheffield Cell network featuring the flow of dihydrogen, electrons, protons, and oxygen.
The Head of the figure above (Figure 2) depicts dihydrogen (double circle) entering MBH (oval) and protons (positive sign) expelled back into the external space of the cell upon hydrolysation. The transport of electrons (negative sign) can be visualised in tandem after the expulsion from MBH and MtrCAB (small rectangle) membrane bound proteins, readily engaging with iron oxide (molecule pictured with 2 irons & 3 oxygens). The removal of oxygen (3O2) from the cell via the perovskite oxide membrane (tail end long rectangle), and the protons via the electrolyte membrane (2x rectangles feature either side of the figure) can be seen eagerly reacting to form water (molecule pictured 1 hydrogen & 2 oxygens) as a byproduct at the tail end of the figure. The curved lines express the continuation of the membranes, which indicates separation between each unit of the cell. The exit point of Fe3+ has not been considered in this figure. Please refer to ‘Sheffield Cell composition’ section for the mechanism of the cell’s interior components for greater detail.
Considerations for Testing the Sheffield Cell
This essay has brought together independent disciplines of biochemical research, which are incorporated into a theoretical chemical cell for the purpose of bioremediation abstraction of iron from iron ore. It has discussed factors that are required for a productive biotechnology model, such as alterations in protein structure and how these alter protein functions in aerotolerance and electron transport. It also gave an overview of the construction of synthetic membranes formed taking advantage of biomaterials, as well as modern techniques for protein positioning within the membrane structure.
Although there has been limited discussion on the practicality of the Sheffield Cell, the following review will give the reader a greater insight into the cell’s fundamentals. In this sense this essay touched on techniques combining the individual components of the cell in forming one functional unit. Currently, there is little evidence regarding the hybridisation of the cell’s subunits, although much research into the single elements remains vibrant within biotech usefulness. For example, the cell’s protein interactions are foreign to one another in natural systems, therefore when choosing the membrane insertion technique precautions should be met to prevent denaturing of the proteins and maximise their efficiency. Furthermore, I mentioned a few methods which would be most suitable, such as the ‘grafting to principle’ technique.
The Sheffield Cell consists of mainly biological components where maintaining equilibrium contributes to increased levels of performance and higher yields. To preserve the cell’s environment, it is necessary to remove iron ions, water molecules, and heat emitted by the cell due to energy transfer. It is also important to be aware that, in order to produce functional proteins with the correct fold and catalytic potential, the expression of MBH and MtrCAB must be facilitated by a complementary host. There are many additional factors which could also be mentioned, such as physiological pH, but they go beyond the scope of this essay.
To conclude, bioremediation has a bright future ahead of it, bringing forward the ‘Green industrial revolution’. Innovation and creativity in the realm of life sciences will allow for a greater reduction in emissions and a stronger global economy due to lower costs and higher output. Technology such as the Sheffield Cell, although an emergent idea, has the potential to bring multiple sectors together for advanced scientific breakthroughs, benefiting society in terms of health, expertise, and living environment.
Equations Relating to the Production of Pure Iron
$$1000gFe*1\frac{mol}{55.845gFe} = 17.9mols\ Fe$$
$$17.9\ mols\ Fe*\frac{{2Fe}_{2}O_{3}}{4Fe\ mols} = 8.95\ mols\ of\ {Fe}_{2}O_{3}$$
(55.845 ×2) + (16 ×3) = 159.69gmol
159.69 × 8.95 = 1429.22 g of iron oxide is needed to produce 1kg of Iron. The reaction will also produce 483.7g of water and require 54.1g of H2 per 1kg of iron produced.
References
Ambrosio-Albalá, P., Upham, P.J., and Gale, W.F. (2023). ‘Normative expectations of government as a policy actor: the case of UK steel industry decarbonisation’, International Journal of Sustainable Energy, 42(1), pp.594–611. Available at: https://doi.org/10.1080/14786451.2023.2217948
Bretschger, O., Obraztsova, A., Sturm, C.A., Chang, I.S., Gorby, Y.A., Reed, S.B., Culley, D.E., Reardon, C.L., Barua, S., Romine, M.F., Zhou, J., Beliaev, A.S., Bouhenni, R., Saffarini, D., Mansfeld, F., Kim, B.-H., Fredrickson, J.K. and Nealson, K.H. (2007) ‘Current Production and Metal Oxide Reduction by Shewanella oneidensis MR-1 Wild Type and Mutants’, Applied and Environmental Microbiology, 73(21), pp.7003–7012. Available at: https://doi.org/10.1128/aem.01087-07
Bruns, N. and Kilbinger, A.F. (eds.). (2016). Bio-inspired polymers. Polymer Chemistry Series, Vol. 22. Cambridge, England: Royal Society of Chemistry.
Edwards, M., White, G., Butt, J., Richardson, D.J. and Clarke, T.A. (2020) ‘The Structure of a Biological Insulated Transmembrane Molecular Wire’, SSRN Electronic Journal, 181(3). Available at: https://doi.org/10.2139/ssrn.3445677
Edwards, M.J., White, G.F., Lockwood, C.W., Lawes, M.C., Martel, A., Harris, G., Scott, D.J., Richardson, D.J., Butt, J.N. and Clarke, T.A. (2018). Structural modeling of an outer membrane electron conduit from a metal-reducing bacterium suggests electron transfer via periplasmic redox partners. Journal of Biological Chemistry, 293(21), pp.8103–8112. Available at: https://doi.org/10.1074/jbc.ra118.001850
Fontecilla-Camps, J.C., Volbeda, A., Cavazza, C. and Nicolet, Y. (2007) ‘Structure/Function Relationships of [NiFe]- and [FeFe]-Hydrogenases’, ChemInform, 38(50). Available at: https://doi.org/10.1002/chin.200750260
Fritsch, J., Scheerer, P., Frielingsdorf, S., Kroschinsky, S., Friedrich, B., Lenz, O., and Spahn, C.M.T. (2011) ‘The crystal structure of an oxygen-tolerant hydrogenase uncovers a novel iron-sulphur centre’, Nature, 479(7372), pp.249–252. Available at: https://doi.org/10.1038/nature10505
Higuchi, Y., Yagi, T., and Yasuoka, N. (1997) ‘Unusual ligand structure in Ni–Fe active center and an additional Mg site in hydrogenase revealed by high resolution X-ray structure analysis’, Structure, 5(12), pp.1671–1680. Available at: https://doi.org/10.1016/s0969-2126(97)00313-4
Knight, L., and Walker, A. (2022) Green Steel. London: UK Parliament POST. Available at: https://post.parliament.uk/research-briefings/post-pn-0672/#:~:text=The%20three%20main%20technology%20pathways,technology%20with%20existing%20steelmaking%20infrastructure (Accessed: 28 August 2024).
Lubitz, W., Ogata, H., Rüdiger, O., and Reijerse, E. (2014) ‘Hydrogenases’, Chemical Reviews, 114(8), pp.4081–4148. Available at: https://doi.org/10.1021/cr4005814
Pathy, A., Nageshwari, K., Ramaraj, R., Pragas Maniam, G., Govindan, N., and Balasubramanian, P. (2022) ‘Biohydrogen production using algae: Potentiality, economics and challenges’, Bioresource Technology, 360, p.127514. Available at: https://doi.org/10.1016/j.biortech.2022.127514
Selyanchyn, O., Selyanchyn, R., and Lyth, S.M. (2020) ‘A Review of Proton Conductivity in Cellulosic Materials’, Frontiers in Energy Research, 8. Available at: https://doi.org/10.3389/fenrg.2020.596164
Shomura, Y., Yoon, K., Nishihara, H., and Higuchi, Y. (2011) ‘Structural basis for a [4Fe-3S] cluster in the oxygen-tolerant membrane-bound [NiFe]-hydrogenase’, Nature (London), 479(7372), pp.253–256. Available at: https://doi.org/10.1038/nature10504
Siekierka, A., Nowicka, J., and Ostrowska, M. (2023) ‘Mechanism of selective transportation of metal ions across chelating membranes in electrodialysis’, Chemical engineering and processing, 189, p.109408. Available at: https://doi.org/10.1016/j.cep.2023.109408.
Sunarso, J., Hashim, S.S., Zhu, N. and Zhou, W. (2017) ‘Perovskite oxides applications in high temperature oxygen separation, solid oxide fuel cell and membrane reactor: A review’, Progress in Energy and Combustion Science, 61, pp.57–77. Available at: https://doi.org/10.1016/j.pecs.2017.03.003
Zheng, Q., Xie, Y., Tan, J., Xu, Z., Luo, P., Wang, T., Liu, Z., Liu, F., Zhang, K., Fang, Z., Zhang, G. and Jin, W. (2022) ‘Coupling of dielectric barrier discharge plasma with oxygen permeable membrane for highly efficient low-temperature permeation’, Journal of Membrane Science, 641, p.119896. Available at: https://doi.org/10.1016/j.memsci.2021.119896
Zhu, X. and Yang, W. (2019) ‘Microstructural and Interfacial Designs of Oxygen‐Permeable Membranes for Oxygen Separation and Reaction–Separation Coupling’, Advanced Materials, 31(50), p.1902547. Available at: https://doi.org/10.1002/adma.201902547
©Daniel Steven Harris. This article is licensed under a Creative Commons Attribution 4.0 International Licence (CC BY).